
Medullary thymic epithelial cells
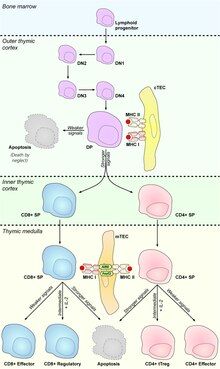
Medullary thymic epithelial cells (mTECs) represent a unique stromal cell population of the thymus which plays an essential role in the establishment of central tolerance. Therefore, mTECs rank among cells relevant for the development of functional mammal immune system.
T cell precursors rise in bone marrow and migrate through the bloodstream to the thymus for further development. During their maturation in the thymus, they undergo a process called V(D)J recombination which conducts the development of T cell receptors (TCRs). The mechanism of this stochastic process enables on one hand the generation of vast repertoire of TCRs, however, on the other hand causes also origin of so called "autoreactive T cells" which recognize self antigens via their TCRs. Autoreactive T cells must be eliminated from the body or skewed into the T Regulatory cells (TRegs) lineage to prevent manifestations of autoimmunity. mTECs possess the ability to deal with these autoreactive clones via mediation of the processes of central tolerance, namely clonal deletion or T regulatory cells selection, respectively.
N.B.: All the below cited references utilized mouse as a model organism.
Self-antigens generation and presentation
[edit]In 1989, two scientific groups came up with the hypothesis that the thymus expresses genes which are in the periphery, strictly expressed by specific tissues (e.g.: Insulin produced by β cells of the pancreas) to subsequently present these so-called "tissue-restricted antigens" (TRAs) from almost all parts of the body to developing T cells in order to test which TCRs recognize self-tissues and can be therefore harmful to the body.[1] It was found, after more than a decade, that this phenomenon is managed specifically by mTECs in the thymus and was named Promiscuous gene expression (PGE).[2]
Autoimmune regulator
[edit]Aire is a protein called autoimmune regulator (Aire) which is also specifically expressed by mTECs.[3] and its expression is completely dependent on NF- kappa B signaling pathway.[4] Aire recognizes target genes of TRAs via specific methylation marks[5][6] and requires about 50 partner molecules for activation of their expression.[7] Moreover, Aire-dependent activation of TRA genes expression is accompanied by formation of DNA double-strand breaks.[8] which probably results in very short lifespan of mTECs between 2–3 days[9]
Mutations of Aire gene in human cause a rare autoimmune disorder called Autoimmune Polyendocrinopathy Candidiasis Ectodermal Distrophy (APECED).,[10][11] which usually manifests in combination with other autoimmune diseases e.g.: diabetes mellitus type 1. Dysfunction of murine Aire gene results in comparable scenario and therefore mouse is used as the model organism for investigation of APECED.
mTECs in numbers
[edit]mTECs as a population are capable to express more than 19000 genes (about 80% of mouse genome) among which approximately 4000 belong to Aire-dependent TRAs. It is important to emphasize that single mTEC expresses about 150 Aire-dependent TRAs and approximately 600 Aire-independent TRAs,[12] indicating that other still unknown PGE regulators exist. Indeed, another protein called Fezf2 was suggested to be the second regulator of PGE.[13]
It was shown that each mTEC expresses stochastically 1-3% of TRA pool.[14] However, more recent studies discovered stable co-expression patterns between TRA genes which are localized in close proximity, suggesting "order in this stochastic process".[15][16]
Tissues protection against autoreactive T cells
[edit]T cell precursors extravasate from the bloodstream in cortico-medullary junction and they first migrate to the thymic cortex, where they undergo construction of TCRs and subsequently a process called T cell positive selection which is mediated by mTEC-related cells: cortical thymic epithelial cells (cTECs). This process verifies, whether newly generated TCRs are functional.[17] About 90% of T cells displays badly rearranged TCRs, they cannot reach the positive selection and they die by neglect in the cortex.[18] The rest starts to express CCR7, which is a receptor for mTEC-generated chemokine CCL21, and migrate after concentration gradient to the thymic medulla to encounter mTECs.[19]
Two modes of central tolerance
[edit]mTECs are not only mediators of PGE and "factories of TRAs". They also express high levels of MHC II and costimulatory molecules CD80/CD86 and rank among efficient antigen-presenting cells (APCs).[2] Moreover, they utilize macroautophagy to load self antigens on MHCII molecules.[20] Thus, mTECs are capable to present self-generated TRAs on their MHC molecules to select potential autoreactive T cells. It was published that mTECs mediate clonal deletion (recessive tolerance), via presentation of TRAs, which leads to the apoptosis of autoreactive T cells,[21][22][23] as well as they are competent to skew autoreactive T cells into TRegs, also through the presentation of TRAs, which then migrate to the periphery to protect tissues against autoreactive T cells that occasionally avoid selection processes in the thymus (dominant tolerance).[24][25]
How mTECs discriminate between these two modes of tolerance? It was shown that prospective TRegs interact with presented TRAs with lower affinity than those which are clonally deleted.[17] Furthermore, it was also revealed that specific TRAs skew autoreactive T cells into TRegs with much higher efficiency than they do in the case of clonal deletion.[26]
Antigen transfer in the thymus
[edit]mTECs form rare population which is composed of approximately 100000 cells per thymus of 2-week-old mice.[27] Thus, there is low probability of encounter between autoreactive T cell and mTEC. Unidirectional antigen transfer from mTECs to thymic dendritic cells (DCs), which itself can't express TRAs, extends the network of TRA presentation, enables TRA processing by different microenvironments and increases the probability of encounter between autoreactive T cell and its appropriate self-antigen.[28][29][30] Moreover, DCs competently induce both recessive and dominant tolerance as well as mTECs.[29]
In contrast, another seminal study reveals that mTECs itself suffice to establish both recessive and dominant tolerance without help of additional APCs.[31]
Development
[edit]Subsets
[edit]mTEC population is not homogenous and basically could be subdivided into more numerous population of mTECs which express low number of MHCII and CD80/CD86, namely mTECsLo and smaller population of mTECsHi which express higher amounts of these molecules.[32] PGE regulator Aire is expressed only by part of mTECsHi.[9] However, this claim does not mean that mTECsLo don't contribute to PGE, mTECsHi, especially that expressing Aire, are just much more efficient in this process.[32]
There is evidence that mTECsLo serve as precursors of mTECsHi in the embryonic thymus[33][34] Nevertheless, situation changes after birth, where only part of mTECsLo pool represents immature mTECsHi reservoir[33] and another part is constituted by mature mTECs which are specialized for expression of chemokine CCL21,[35] discussed above. Further subset of mTECsLo pool is formed by terminally differentiated cells called Post- Aire mTECs which already downregulated the expression of Aire, MHCII and CD80/CD86.[36]
mTECs can develop into Thymic mimetic cells, which combine the mTEC identity with lineage specific transcription factors. These cells exhibit the phenotype of differentiated peripheral cells and produce their corresponding TRAs. The most famous example is Hassall's corpuscles.[37]
Progenitor cells
[edit]TECs (mTECs and cTECs) originate from the third pharyngeal pouch which is a product of endoderm.[38] Their common origin points to the fact that both mTECs and cTECs rise from one bipotent progenitor. This notion was confirmed by several studies of embryonic thymus.[39][40] and was further developed by finding that these bipotent progenitors express cTEC markers.[41][42] Nevertheless, another sources document existence of mTEC unipotent progenitors that express claudin 3 and 4 (Cld3/4).[43][44] These two opposite findings were interfaced by observation of unipotent mTEC progenitors in the postnatal thymus that previously expressed cTEC markers and concurrently express Cld3/4.[45] On the other hand, several other studies describe appearance of bipotent progenitors in postnatal thymus.[46][47][48][49] Thus, embryonic as well as postnatal thymus might shelter both bipotent TEC or unipotent mTEC progenitors.
Similarly to Aire expression, mTECs development is highly dependent on NF- kappa B signaling pathway.[50]
References
[edit]- ^ Linsk R, Gottesman M, Pernis B (October 1989). "Are tissues a patch quilt of ectopic gene expression?". Science. 246 (4927): 261. Bibcode:1989Sci...246..261L. doi:10.1126/science.2799388. PMID 2799388.
- ^ a b Derbinski J, Schulte A, Kyewski B, Klein L (November 2001). "Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self". Nature Immunology. 2 (11): 1032–9. doi:10.1038/ni723. PMID 11600886. S2CID 20155713.
- ^ Anderson MS, Venanzi ES, Klein L, Chen Z, Berzins SP, Turley SJ, von Boehmer H, Bronson R, Dierich A, Benoist C, Mathis D (November 2002). "Projection of an immunological self shadow within the thymus by the aire protein". Science. 298 (5597): 1395–401. Bibcode:2002Sci...298.1395A. doi:10.1126/science.1075958. PMID 12376594. S2CID 13989491.
- ^ Haljasorg U, Bichele R, Saare M, Guha M, Maslovskaja J, Kõnd K, Remm A, Pihlap M, Tomson L, Kisand K, Laan M, Peterson P (December 2015). "A highly conserved NF-κB-responsive enhancer is critical for thymic expression of Aire in mice". European Journal of Immunology. 45 (12): 3246–56. doi:10.1002/eji.201545928. PMID 26364592.
- ^ Org T, Chignola F, Hetényi C, Gaetani M, Rebane A, Liiv I, Maran U, Mollica L, Bottomley MJ, Musco G, Peterson P (April 2008). "The autoimmune regulator PHD finger binds to non-methylated histone H3K4 to activate gene expression". EMBO Reports. 9 (4): 370–6. doi:10.1038/embor.2008.11. PMC 2261226. PMID 18292755.
- ^ Koh AS, Kuo AJ, Park SY, Cheung P, Abramson J, Bua D, Carney D, Shoelson SE, Gozani O, Kingston RE, Benoist C, Mathis D (October 2008). "Aire employs a histone-binding module to mediate immunological tolerance, linking chromatin regulation with organ-specific autoimmunity". Proceedings of the National Academy of Sciences of the United States of America. 105 (41): 15878–83. Bibcode:2008PNAS..10515878K. doi:10.1073/pnas.0808470105. PMC 2572939. PMID 18840680.
- ^ Abramson J, Giraud M, Benoist C, Mathis D (January 2010). "Aire's partners in the molecular control of immunological tolerance". Cell. 140 (1): 123–35. doi:10.1016/j.cell.2009.12.030. PMID 20085707.
- ^ Guha M, Saare M, Maslovskaja J, Kisand K, Liiv I, Haljasorg U, Tasa T, Metspalu A, Milani L, Peterson P (April 2017). "DNA breaks and chromatin structural changes enhance the transcription of autoimmune regulator target genes". The Journal of Biological Chemistry. 292 (16): 6542–6554. doi:10.1074/jbc.m116.764704. PMC 5399106. PMID 28242760.
- ^ a b Gray D, Abramson J, Benoist C, Mathis D (October 2007). "Proliferative arrest and rapid turnover of thymic epithelial cells expressing Aire". The Journal of Experimental Medicine. 204 (11): 2521–8. doi:10.1084/jem.20070795. PMC 2118482. PMID 17908938.
- ^ Nagamine K, Peterson P, Scott HS, Kudoh J, Minoshima S, Heino M, Krohn KJ, Lalioti MD, Mullis PE, Antonarakis SE, Kawasaki K, Asakawa S, Ito F, Shimizu N (December 1997). "Positional cloning of the APECED gene". Nature Genetics. 17 (4): 393–8. doi:10.1038/ng1297-393. PMID 9398839. S2CID 1583134.
- ^ Kisand K, Peterson P (July 2015). "Autoimmune polyendocrinopathy candidiasis ectodermal dystrophy". Journal of Clinical Immunology. 35 (5): 463–78. doi:10.1007/s10875-015-0176-y. PMID 26141571. S2CID 8080647.
- ^ Sansom SN, Shikama-Dorn N, Zhanybekova S, Nusspaumer G, Macaulay IC, Deadman ME, Heger A, Ponting CP, Holländer GA (December 2014). "Population and single-cell genomics reveal the Aire dependency, relief from Polycomb silencing, and distribution of self-antigen expression in thymic epithelia". Genome Research. 24 (12): 1918–31. doi:10.1101/gr.171645.113. PMC 4248310. PMID 25224068.
- ^ Takaba H, Morishita Y, Tomofuji Y, Danks L, Nitta T, Komatsu N, Kodama T, Takayanagi H (November 2015). "Fezf2 Orchestrates a Thymic Program of Self-Antigen Expression for Immune Tolerance". Cell. 163 (4): 975–87. doi:10.1016/j.cell.2015.10.013. PMID 26544942.
- ^ Derbinski J, Pinto S, Rösch S, Hexel K, Kyewski B (January 2008). "Promiscuous gene expression patterns in single medullary thymic epithelial cells argue for a stochastic mechanism". Proceedings of the National Academy of Sciences of the United States of America. 105 (2): 657–62. Bibcode:2008PNAS..105..657D. doi:10.1073/pnas.0707486105. PMC 2206592. PMID 18180458.
- ^ Brennecke P, Reyes A, Pinto S, Rattay K, Nguyen M, Küchler R, Huber W, Kyewski B, Steinmetz LM (September 2015). "Single-cell transcriptome analysis reveals coordinated ectopic gene-expression patterns in medullary thymic epithelial cells". Nature Immunology. 16 (9): 933–41. doi:10.1038/ni.3246. PMC 4675844. PMID 26237553.
- ^ Rattay K, Meyer HV, Herrmann C, Brors B, Kyewski B (February 2016). "Evolutionary conserved gene co-expression drives generation of self-antigen diversity in medullary thymic epithelial cells". Journal of Autoimmunity. 67: 65–75. doi:10.1016/j.jaut.2015.10.001. PMID 26481130.
- ^ a b Klein L, Kyewski B, Allen PM, Hogquist KA (June 2014). "Positive and negative selection of the T cell repertoire: what thymocytes see (and don't see)". Nature Reviews. Immunology. 14 (6): 377–91. doi:10.1038/nri3667. PMC 4757912. PMID 24830344.
- ^ Palmer E (May 2003). "Negative selection--clearing out the bad apples from the T-cell repertoire". Nature Reviews. Immunology. 3 (5): 383–91. doi:10.1038/nri1085. PMID 12766760. S2CID 28321309.
- ^ Kurobe H, Liu C, Ueno T, Saito F, Ohigashi I, Seach N, Arakaki R, Hayashi Y, Kitagawa T, Lipp M, Boyd RL, Takahama Y (February 2006). "CCR7-dependent cortex-to-medulla migration of positively selected thymocytes is essential for establishing central tolerance". Immunity. 24 (2): 165–77. doi:10.1016/j.immuni.2005.12.011. PMID 16473829.
- ^ Aichinger M, Wu C, Nedjic J, Klein L (February 2013). "Macroautophagy substrates are loaded onto MHC class II of medullary thymic epithelial cells for central tolerance". The Journal of Experimental Medicine. 210 (2): 287–300. doi:10.1084/jem.20122149. PMC 3570095. PMID 23382543.
- ^ Liston A, Lesage S, Wilson J, Peltonen L, Goodnow CC (April 2003). "Aire regulates negative selection of organ-specific T cells". Nature Immunology. 4 (4): 350–4. doi:10.1038/ni906. PMID 12612579. S2CID 4561402.
- ^ Anderson MS, Venanzi ES, Chen Z, Berzins SP, Benoist C, Mathis D (August 2005). "The cellular mechanism of Aire control of T cell tolerance". Immunity. 23 (2): 227–39. doi:10.1016/j.immuni.2005.07.005. PMID 16111640.
- ^ Taniguchi RT, DeVoss JJ, Moon JJ, Sidney J, Sette A, Jenkins MK, Anderson MS (May 2012). "Detection of an autoreactive T-cell population within the polyclonal repertoire that undergoes distinct autoimmune regulator (Aire)-mediated selection". Proceedings of the National Academy of Sciences of the United States of America. 109 (20): 7847–52. Bibcode:2012PNAS..109.7847T. doi:10.1073/pnas.1120607109. PMC 3356674. PMID 22552229.
- ^ Aschenbrenner K, D'Cruz LM, Vollmann EH, Hinterberger M, Emmerich J, Swee LK, Rolink A, Klein L (April 2007). "Selection of Foxp3+ regulatory T cells specific for self antigen expressed and presented by Aire+ medullary thymic epithelial cells". Nature Immunology. 8 (4): 351–8. doi:10.1038/ni1444. PMID 17322887. S2CID 21052344.
- ^ Malchow S, Leventhal DS, Nishi S, Fischer BI, Shen L, Paner GP, Amit AS, Kang C, Geddes JE, Allison JP, Socci ND, Savage PA (March 2013). "Aire-dependent thymic development of tumor-associated regulatory T cells". Science. 339 (6124): 1219–24. Bibcode:2013Sci...339.1219M. doi:10.1126/science.1233913. PMC 3622085. PMID 23471412.
- ^ Malchow S, Leventhal DS, Lee V, Nishi S, Socci ND, Savage PA (May 2016). "Aire Enforces Immune Tolerance by Directing Autoreactive T Cells into the Regulatory T Cell Lineage". Immunity. 44 (5): 1102–13. doi:10.1016/j.immuni.2016.02.009. PMC 4871732. PMID 27130899.
- ^ Klein L (August 2009). "Dead man walking: how thymocytes scan the medulla". Nature Immunology. 10 (8): 809–11. doi:10.1038/ni0809-809. PMID 19621041. S2CID 28668641.
- ^ Koble C, Kyewski B (July 2009). "The thymic medulla: a unique microenvironment for intercellular self-antigen transfer". The Journal of Experimental Medicine. 206 (7): 1505–13. doi:10.1084/jem.20082449. PMC 2715082. PMID 19564355.
- ^ a b Perry JS, Lio CJ, Kau AL, Nutsch K, Yang Z, Gordon JI, Murphy KM, Hsieh CS (September 2014). "Distinct contributions of Aire and antigen-presenting-cell subsets to the generation of self-tolerance in the thymus". Immunity. 41 (3): 414–426. doi:10.1016/j.immuni.2014.08.007. PMC 4175925. PMID 25220213.
- ^ Kroger CJ, Spidale NA, Wang B, Tisch R (January 2017). "Thymic Dendritic Cell Subsets Display Distinct Efficiencies and Mechanisms of Intercellular MHC Transfer". Journal of Immunology. 198 (1): 249–256. doi:10.4049/jimmunol.1601516. PMC 5173434. PMID 27895179.
- ^ Hinterberger M, Aichinger M, Prazeres da Costa O, Voehringer D, Hoffmann R, Klein L (June 2010). "Autonomous role of medullary thymic epithelial cells in central CD4(+) T cell tolerance". Nature Immunology. 11 (6): 512–9. doi:10.1038/ni.1874. PMID 20431619. S2CID 33154019.
- ^ a b Derbinski J, Gäbler J, Brors B, Tierling S, Jonnakuty S, Hergenhahn M, Peltonen L, Walter J, Kyewski B (July 2005). "Promiscuous gene expression in thymic epithelial cells is regulated at multiple levels". The Journal of Experimental Medicine. 202 (1): 33–45. doi:10.1084/jem.20050471. PMC 2212909. PMID 15983066.
- ^ a b Gäbler J, Arnold J, Kyewski B (December 2007). "Promiscuous gene expression and the developmental dynamics of medullary thymic epithelial cells". European Journal of Immunology. 37 (12): 3363–72. doi:10.1002/eji.200737131. PMID 18000951.
- ^ Rossi SW, Kim MY, Leibbrandt A, Parnell SM, Jenkinson WE, Glanville SH, McConnell FM, Scott HS, Penninger JM, Jenkinson EJ, Lane PJ, Anderson G (June 2007). "RANK signals from CD4(+)3(-) inducer cells regulate development of Aire-expressing epithelial cells in the thymic medulla". The Journal of Experimental Medicine. 204 (6): 1267–72. doi:10.1084/jem.20062497. PMC 2118623. PMID 17502664.
- ^ Lkhagvasuren E, Sakata M, Ohigashi I, Takahama Y (May 2013). "Lymphotoxin β receptor regulates the development of CCL21-expressing subset of postnatal medullary thymic epithelial cells". Journal of Immunology. 190 (10): 5110–7. doi:10.4049/jimmunol.1203203. PMID 23585674.
- ^ Metzger TC, Khan IS, Gardner JM, Mouchess ML, Johannes KP, Krawisz AK, Skrzypczynska KM, Anderson MS (October 2013). "Lineage tracing and cell ablation identify a post-Aire-expressing thymic epithelial cell population". Cell Reports. 5 (1): 166–79. doi:10.1016/j.celrep.2013.08.038. PMC 3820422. PMID 24095736.
- ^ Michelson, Daniel A.; Mathis, Diane (October 2022). "Thymic mimetic cells: tolerogenic masqueraders". Trends in Immunology. 43 (10): 782–791. doi:10.1016/j.it.2022.07.010. ISSN 1471-4906. PMC 9509455. PMID 36008259.
- ^ Gordon J, Wilson VA, Blair NF, Sheridan J, Farley A, Wilson L, Manley NR, Blackburn CC (May 2004). "Functional evidence for a single endodermal origin for the thymic epithelium". Nature Immunology. 5 (5): 546–53. doi:10.1038/ni1064. PMID 15098031. S2CID 10282524.
- ^ Bleul CC, Corbeaux T, Reuter A, Fisch P, Mönting JS, Boehm T (June 2006). "Formation of a functional thymus initiated by a postnatal epithelial progenitor cell". Nature. 441 (7096): 992–6. Bibcode:2006Natur.441..992B. doi:10.1038/nature04850. PMID 16791198. S2CID 4396922.
- ^ Rossi SW, Jenkinson WE, Anderson G, Jenkinson EJ (June 2006). "Clonal analysis reveals a common progenitor for thymic cortical and medullary epithelium". Nature. 441 (7096): 988–91. Bibcode:2006Natur.441..988R. doi:10.1038/nature04813. PMID 16791197. S2CID 2358330.
- ^ Baik S, Jenkinson EJ, Lane PJ, Anderson G, Jenkinson WE (March 2013). "Generation of both cortical and Aire(+) medullary thymic epithelial compartments from CD205(+) progenitors". European Journal of Immunology. 43 (3): 589–94. doi:10.1002/eji.201243209. PMC 3960635. PMID 23299414.
- ^ Ohigashi I, Zuklys S, Sakata M, Mayer CE, Zhanybekova S, Murata S, Tanaka K, Holländer GA, Takahama Y (June 2013). "Aire-expressing thymic medullary epithelial cells originate from β5t-expressing progenitor cells". Proceedings of the National Academy of Sciences of the United States of America. 110 (24): 9885–90. Bibcode:2013PNAS..110.9885O. doi:10.1073/pnas.1301799110. PMC 3683726. PMID 23720310.
- ^ Hamazaki Y, Fujita H, Kobayashi T, Choi Y, Scott HS, Matsumoto M, Minato N (March 2007). "Medullary thymic epithelial cells expressing Aire represent a unique lineage derived from cells expressing claudin". Nature Immunology. 8 (3): 304–11. doi:10.1038/ni1438. PMID 17277780. S2CID 7775843.
- ^ Sekai M, Hamazaki Y, Minato N (November 2014). "Medullary thymic epithelial stem cells maintain a functional thymus to ensure lifelong central T cell tolerance". Immunity. 41 (5): 753–61. doi:10.1016/j.immuni.2014.10.011. PMID 25464854.
- ^ Ohigashi I, Zuklys S, Sakata M, Mayer CE, Hamazaki Y, Minato N, Hollander GA, Takahama Y (November 2015). "Adult Thymic Medullary Epithelium Is Maintained and Regenerated by Lineage-Restricted Cells Rather Than Bipotent Progenitors". Cell Reports. 13 (7): 1432–1443. doi:10.1016/j.celrep.2015.10.012. hdl:2433/216325. PMID 26549457.
- ^ Ucar A, Ucar O, Klug P, Matt S, Brunk F, Hofmann TG, Kyewski B (August 2014). "Adult thymus contains FoxN1(-) epithelial stem cells that are bipotent for medullary and cortical thymic epithelial lineages". Immunity. 41 (2): 257–69. doi:10.1016/j.immuni.2014.07.005. PMC 4148705. PMID 25148026.
- ^ Wong K, Lister NL, Barsanti M, Lim JM, Hammett MV, Khong DM, Siatskas C, Gray DH, Boyd RL, Chidgey AP (August 2014). "Multilineage potential and self-renewal define an epithelial progenitor cell population in the adult thymus". Cell Reports. 8 (4): 1198–209. doi:10.1016/j.celrep.2014.07.029. PMID 25131206.
- ^ Ulyanchenko S, O'Neill KE, Medley T, Farley AM, Vaidya HJ, Cook AM, Blair NF, Blackburn CC (March 2016). "Identification of a Bipotent Epithelial Progenitor Population in the Adult Thymus". Cell Reports. 14 (12): 2819–32. doi:10.1016/j.celrep.2016.02.080. PMC 4819909. PMID 26997270.
- ^ Meireles C, Ribeiro AR, Pinto RD, Leitão C, Rodrigues PM, Alves NL (June 2017). "Thymic crosstalk restrains the pool of cortical thymic epithelial cells with progenitor properties". European Journal of Immunology. 47 (6): 958–969. doi:10.1002/eji.201746922. hdl:10216/111812. PMID 28318017.
- ^ van Delft MA, Huitema LF, Tas SW (May 2015). "The contribution of NF-κB signalling to immune regulation and tolerance". European Journal of Clinical Investigation. 45 (5): 529–39. doi:10.1111/eci.12430. PMID 25735405.